ЛЕКЦИЯ
№7.
ОТДЕЛ БУРЫЕ ВОДОРОСЛИ - PHAEOPHYTA
Общая харатеристика. Строение
слоевища и клеток. Размножение и
циклы развития.
Класс Изогенератные.
Класс Гетерогенератные.
Класс Циклоспоровые.
Общая характеристика. Строение слоевища и
клеток.
Размножение и циклы развития.
Бурых водорослей
насчитывает 1500 видов, 240 родов, 10 – 14 порядков.
Это, за
исключением представителей трех родов, морские водоросли. Особенно богато
представлены они в холодных водах северного и южного полушария до глубины 15
м., хотя некоторые виды саргассума и ламинарий найдены на глубине 200 м. Все
представители – многоклеточные водоросли, размеры которых варьируют от нескольких
мм до нескольких десятков м.: 60, 100, 200, 400 (макроцистис). Среди бурых нет
ни одноклеточных, ни колониальных форм, ни простых неразветвленных нитей. По
внешнему виду это ветвистые кустики и мешочки, корочки, ленты, пластинки,
которые у высокоорганизованных водорослей расчленены на стеблевидные и
листовидные структуры (саргассум, ламинария, лессония). Все бурые водоросли
ведут прикрепленный образ жизни с помощью ризоидов или дисковидных разрастаний.
Талломы нарастают за счет верхушек, интеркалярных клеток или диффузно. Они
могут быть однолетними и многолетними. В многорядных слоевищах
высокоорганизованных водорослей наблюдается специализация клеток.
Анатомическое строение в данном случае сложное, т.к. наблюдается ложно - или
даже истинно-тканевое строение. В их теле могут присутствовать 4 типа тканей –
ассимиляционные, запасающие, проводящие, покровные.
Талломы окрашены
в разные оттенки желтовато-бурого цвета, вследствие содержания в их клетках
хлорофиллов а и с, α- и β-каротинов и бурых ксантофиллов (фукоксантина,
антераксантин, зеаксан-тина, виолаксантина).
Клетки покрыты
целлюлозной оболочкой, которая по своим свойствам отличается от целлюлозы
высших растений, и поэтому ее иногда называют альгулезой. Наружный слой
оболочки представлен пектиновыми веществами, состоящие в основном из альгиновой
кислоты. Набухая в воде, они образуют слизистую консистенцию, которая
предохраняет водоросли от высыхания или механических повреждений во время
отлива. В клетках имеется одно ядро, одно или несколько вакуолей с клеточным соком
щелочной природы и хроматофоры, которые называются феопласты. Они могут быть в виде зерен, дисков, пластин (в
последнем случае с пиреноидами). У эктокарповых, кутлериевых пиреноиды имеются,
у ламинариевых встречаются крайне редко, у диктиотовых, тилоптеридовых – они
отсутствуют, а у сфацеляриевых и фукусовых имеются лишь в хлоропластах гамет.
Пиреноиды очень мелкие и выступают с поверхности хроматофора в цитоплазму (рис.21.)

Рис. 21. Схема строения клетки бурых водорослей:
кс
– клеточная стенка; м – митохондрия; эс – фрагмент канала эндоплазматической
сети; д – диктиосомы; в – вакуоль; я – ядро;
яо
– ядерная оболочка (в перинуклеарном пространстве заключен хлоропласт); х –
хлоропласт; т – трехтилакоидная ламелла; п – пиреноид с обкладкой; г –
кольцевидный генофор, перерезанный поперек
Они окружены
сложной системой мембран. Их четыре. Ламелы сгруппированы по три
(трехтилакоидные). Запасной углевод, полисахарид ламинарии,
откладывается в цитоплазме.
Запасными продуктами являются также шестиатомный сахароспирт маннит и жиры.
В цитоплазме находятся еще мелкие вакуоли – физоды. Их содержимое называется фукозаном
и включает большое количество дубильных веществ в виде флороглицина и других
полифенолов.
Размножение
вегетативное, бесполое и половое. Вегетативное происходит кусочками случайно
оторвавшегося таллома (фрагментация), который не способен прикрепляться к
грунту без ризоидальных структур (у них не формируются ни спорангии, ни
гаметангии). У некоторых образуются выводковые почки, легко отламывающиеся с
образованием новых талломов (сфацелярия). Бесполое размножение у большинства
бурых водорослей происходит посредством гаплоидных зооспор или неподвижных
апланоспор (тетраспор и моноспор), которые образуются в одногнездных
спорангиях после редукционного деления на диплоидных растениях (спорофитах).
Монадные клетки (зооспоры и гаметы) имеют глазок и жгутики. Жгутики в числе
двух, гетероконтные и гетероморфны, перистый жгутик направлен вперед. У
диктиотовых он один, т.к., задний не развит.
Половой процесс
изо-, редко гетеро-(гиффордия) и оогамный. Изо- и гетерогаметы образуются в
многогнездных или многокамерных гаметангиях, по одному в каждом. Иногда
изогаметы могут вести себя как зооспоры, в таком случае гаметангии называют
спорангиями. В оогониях и антеридиях образуется по одной гамете (за исключением
представителей порядка фукусовых). Яйцеклетка оплодотворяется всегда вне
оогония, а зигота прорастает без периода покоя.
У всех бурых
водорослей (кроме циклоспоровых) наблюдается смена поколений – генераций: у
одних изоморфная, у других гетероморфная. На основе особенностей циклов
развития отдел бурых водорослей делится на следующие классы.
Класс Изогенератные -Isogeneratae
Представители
эктокарпус, диктиота, падина, сфацелярия, кутлерия. Смена поколений изоморфная,
но строгое чередование форм развития может и отсутствовать, т.к., из спор
порой развиваются не только гаметофиты, но также спорофиты и гаметоспорофиты.
Порядок эктокарповые – Ectocarpales
Семейство
эктокарповые – Ectocarpaceae
Род эктокарпус – Ectocarpus
Это одни из
примитивных прикрепленных бурых водорослей, распространенных во всех морях, а
особенно холодных. Таллом – желтовато-бурый кустик нескольких см. (30–60) с
пластинчатыми или лентовидными хроматофорами, (рис.22). Эктокарпус состоит из
стелющихся по субстрату нитей, от которых отходят вертикальные однорядные
ветвящиеся нити. Стелющиеся нити имеют верхушечный рост (их считают стелющимися
ризоидами), а вертикальные - диффузный. Крупные вертикальные ветви в основании
бывают покрыты корой из ризоидов. Нити слоевища сужаются к концам и на вершине
оканчиваются длинными бесцветными клетками. Вегетативное размножение у
эктокарповых происходит обрывками нитей.
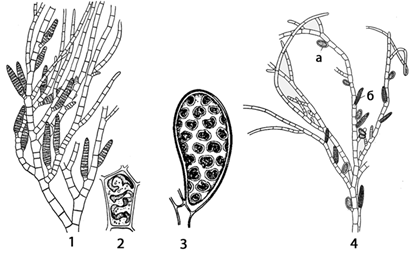
Рис. 22. Ectocarpus: 1 – внешний вид
таллома с многогнездными спорангиями (гаметангиями); 2 – клетка таллома при
большом увеличении; 3 – одногнездный спорангий; 4 – спорофит с одногнездными
(а) и многогнездными (нейтральными, б) спорангиями
На диплоидном
спорофите в одногнездных спорангиях, представляющих собой конечные клетки
боковых ветвей, происходит редукционное деление ядра. При этом образуются
гаплоидные двужгутиковые зооспоры. Они вырастают в гаплоидные растения –
гаметофиты, которые по внешнему виду и размеру не отличаются от спорофита. На
гаметофитах в многогнездных гаметангиях (спорангиях) формируются изогаметы.
Внешне они одинаковы, но отличаются подвижностью. Те, которые быстро теряют
подвижность, являются женскими гаметами. Они выделяют сильно пахучее, летучее
вещество эктокарпен (углеводород), привлекающий мужские гаметы. Одна из них
производит оплодотворение. Зигота без периода покоя прорастает в диплоидное
бесполое растение – спорофит. Иногда гаметы прорастают партеногенети чески
(без оплодотворения) и тогда появляются растения, подобные спорофитам или
гаметофитам.
Цикл
развития Ectocarpus

Порядок диктиотовые - Dictyotales
Семейство
диктиотовые - Dictyotaceae
Род диктиота – Dictyota
Рост апикальный,
дихотомическое ветвление, бесполое размножение с помощью тетраспор, половой
процесс оогамный, растения двудомные, сперматозоиды одножгутиковые, изоморфная
смена поколений.
Диктиота
дихотомическая распространена в Атлантическом океане, по берегам Европы и в
Черном море в местах с движением воды. Таллом лентовидный, дихотомически
ветвящийся в одной плоскости до 20 см. Органом прикрепления служит
цилиндрический ризом – стелющийся побег с ризоидами (рис.23.)

Рис. 23. Dictyota: 1 – внешний вид таллома;
2
– срез спорофита диктиоты с тетраспорангиями; 3 – срез гаметофита
диктиоты
с сорусом оогониев; 4 – то же с сорусом антеридиев
Вершина каждой
ветви заканчивается одной крупной клеткой, за счет чего происходит ее
нарастание. В центре располагается один слой крупных бесцветных клеток,
окруженный снаружи корой из одного слоя мелких интенсивно окрашенных клеток.
Из поверхностных клеток спорофита развиваются крупные одногнездные
тетраспорангии. Они располагаются сорусами. После редукционного деления в них
образуется по четыре гаплоидные неподвижных тетраспоры. Они прорастают в
мужские и женские гаметофиты, внешне похожие на спорофит. На половой генерации
гаметангии собраны группами – сорусами. На
мужских растениях сорус состоит из нескольких многокамерных антеридиев
(антеридии делятся горизонтальными и вертикальными перегородками на 150-1500
камер). В каждой камере образуется один одножгутиковый сперматозоид
(антерозоид). На женских растениях сорусы состоят из нескольких одноклеточных
оогониев, содержащих по одной яйцеклетке. При созревании они вываливаются из
оогониев в воду, где и происходит оплодотворение. Половой процесс оогамный.
Зигота прорастает в спорофит (тетраспорофит) без периода покоя. Т.о., смена
поколений изоморфная.
Цикл
развития Dictyota dichotoma

Класс гетерогенератные -Нeterogeneratae
Представители: Laminaria, Lessonia, Macrocystis, Алария
В жизненном цикле
у представителей происходит чередование сильно отличающихся поколений
(генераций). Спорофит и гаметофит отличаются по форме, величине, внутренней
дифференцировке. Спорофит крупный, гаметофит микроскопический, называемый
заростком.
Порядок ламинариевые - Laminariales
Семейство
ламинариевые - Laminariaceae
Род – Laminaria
Ламинария
(морская капуста) обитает в северных морях. Таллом макроскопический (60 м.) и
расчленен на ризоиды, мощный черешок и листовидную пластинку (рис.24). Черешок
имеет сложное строение и состоит из трех частей. Это кора, клетки которой
содержат хроматофоры и образуют ассимиляционную ткань.

Рис. 24. Морские водоросли порядка Laminariales: 1 – Laminaria saccharina; 2 – Laminaria digitata;
3
– Nereocystis; 4 – Alaria; 5 – Macrocystis
Наружные ее
клетки мелкоклеточные, а внутренние – крупноклеточные. Далее идее
крупноклеточный промежуточный слой.
Центр черешка
занят сердцевиной из длинных неплотно сплетенных нитей-перекладин. Одни из них
идут поперек слоевища, другие ориентированы вдоль оси. В поперечных
перегородках этих последних клеток содержатся многочисленные поры – ситовидные
пластинки. Это проводящие элементы, обеспечивающие транспортировку веществ в
пределах слоевища. В пластинках сердцевинного слоя нет, но они трехслойны: центральные
клетки крупные, а наружные — мелкие.
На границе между
черешком и листом находится интеркалярная зона роста, за счет которой растет в
длину и лист, и стеблеподобный орган. В толщину же рост черешка происходит за
счет деления клеток коры. Слоевища ламинариевых обладают наибольшей скоростью
роста (более 10 см. и сутки), но в верхних концах пластинки происходят постоянные
процессы разрушения. Таллом многолетний, но листоподобная часть периодически
полностью разрушается и отмирает. Это происходит после завершения выхода
спор. Одновременно с выходом спор
начинается закладка новой пластины, поэтому иногда можно наблюдать цепочку из
трех – четырех старых пластин.
Перед сбрасыванием из периферических клеток листьев
диплоидного спорофита образуются сорусы (группы) одноклеточных эллиптических
зооспорангиев, чередующихся с парафизами. Этот процесс начинается с вершины.
После редукционного деления в каждом спорангии образуется равное число спор,
дающих впоследствии мужские и женские гаметофиты. Через отверстие на верхушке
зооспоры выплывают в воду. Они активны от нескольких минут до двух часов.
Встретив твердую поверхность, зооспора прикрепляется передним жгутиком, второй
– теряется. Если за время активного движения она не найдет место для
прикрепления, то наступит неминуемая гибель. Зооспоры прорастают в
микроскопические мужские и женские заростки – гаметофиты, которые различаются
морфологически (мужские мельче и разветвленнее). Они однорядные, несколько
клеточные, но разветвленные. Выросты мужских заростков превращаются в одноклеточные
антеридии где образуется по одному сперматозоиду. В оогоний превращается любая
клетка женского заростка, но больше частью – конечные клетки ветвей. В оогонии
– одна яйцеклетка. Она выталкивается на верхушку оогония (но не отделяется),
где ее оплодотворяет подплывший сперматозоид. Зигота без периода покоя
трогается в рост в нитчатый таллом, который вскоре приступает к дифференцировке
и превращается во взрослый, сложно рассеченный спорофит. Т.о., на месте одного
многоклеточного женского гаметофита развивается несколько спорофитов
У ламинариевых
налицо чередование двух гетероморфных поколений: сложно рассеченного,
дифференцированного, макроскопического спорофита и примитивно устроенного,
несколько клеточного, микроскопического гаметофита.
Цикл
развития Laminaria

Класс Циклоспоровые -Сyclosporeae
Представители: Fucus, Sargassum, Cystoseria, Ascophyllum.
Этот класс
представлен одним порядком фукусовые. У представителей сложно
дифференцированный таллом с верхушечным ростом, отсутствие бесполого размножения,
отсутствие двух самостоятельно растущих форм развития (скрытое чередование
поколений, т.к., гаметофиты развиваются в слоевище спорофита в виде слоя,
выстилающего скафидий) и оогамный половой процесс. Половые органы располагаются
в углублениях таллома – концептакулах
(скафидиях). Мейоз происходит перед образованием гамет.
Порядок фукусовые – Fucales
Семейство
фукусовые - Fucaceae
Род фукус – Fucus
Фукус (ф.
пузырчатый, ф. зубчатый) распространен в северных морях. Таллом узкий, плоский,
ремневидный, дихотомически ветвящийся до 1м в длину со средней жилкой. Состоит из сердцевинной и коровой части. В нижней
части таллом переходит в черешок, который прикрепляется к субстрату расширенным
основанием – диском. У некоторых видов по бокам от срединной жилки имеются
вздутия, заполненные воздухом – воздушные
пузыри, что поддерживает вертикальное положение. Растут ветви за счет одной
верхушечной клетки, расположенных на концах.
Размножение оогамное. При размножении концы таллома вздуваются и принимают
желто-оранжевую окраску, превращаясь в рецептакулы. В них погружены
многочисленные концептакулы (скафидии). Это
округлое, грушевидное углубление в ткани рецептакула, сообщающееся с наружной
средой через узкое отверстие. Выстилающий слой скафидия является гаметофитом.
Он диплоидный и формируется из одной клетки. Внутри от стенок скафидия
отрастают многочисленные волоски – парафизы,
иногда высовывающиеся из отверстия в виде хохолка. В оогониях женского
скафидия после редукционного деления образуется 8 яйцеклеток, в антеридияю
мужского скафидия – 64 сперматозоида.
Растения могут быть как однодомными, так и двудомными. Иногда в одном скафидии
развиваются и антеридии и оогонии. Развивающаяся слизь выталкивает половые
клетки из скафидия. Яйца выделяют летучее вещество фукосерратеном, привлекающее
сперматозоиды. После слияния половых клеток образуется зигота, немедленно
прорастающая в диплоидный таллом – спорофит. Т.о., в жизненном цикле
циклоспоровых нет смены генераций (или есть скрытое поколение в виде выстилающего
слоя скафидия). Присутствует стадия гаметофита, но особь - всегда диплонт, т.к.
мейоз происходит лишь перед образованием гамет. Таким образом, для фукуса характерен
диплонтный цикл развития.

Рис. 25. Строение таллома и размножение у Fucus: a – общий вид таллома; б – разрез женского
скафидия;
в
– разрез мужского скафидия; г – оогоний;
д, е – выход яйцеклеток из оогония; ж – веточка
с
антеридиями; з – выход сперматозоидов из антеридиев; и – сперматозоид
У бурых
водорослей наблюдается смена поколений – генераций: у одних изоморфная, у
других гетероморфная. Однако деление на изогенератные и гетерогенератные очень
условно, поскольку в обоих классах существуют представители с противоположным
типом смены форм развития. Поэтому многие считают целесообразной следующую
классификацию:
Цикл развития Fucus

Большое спасибо
ОтветитьУдалитьСпасибо 👍
ОтветитьУдалить👍
ОтветитьУдалить